Какво е хроматин? Хроматинови функции
Генетичният материал на еукариотните организми има много сложна организация. ДНК молекули, намиращи се в клетъчното ядро, са част от специално многокомпонентно вещество - хроматин.
Дефиниция на концепцията
Хроматинът е материал на клетъчното ядро, съдържащо наследствена информация, което е сложен функционален ДНК комплекс със структурни протеини и други елементи, които осигуряват опаковането, съхранението и реализацията на кариотичния геном. В опростена интерпретация, това е веществото, което съставлява хромозомите. Терминът идва от гръцкия "хром" - цвят, боя.
Концепцията е въведена от Флеминг още през 1880 г., но все още има дебати за това какво е хроматин от гледна точка на неговия биохимичен състав. Несигурността засяга малка част от компонентите, които не участват в структурирането на генетични молекули (някои ензими и рибонуклеинови киселини).
В електронна снимка на интерфазното ядро хроматинът се визуализира като множество петна от тъмна материя, които могат да бъдат малки и разпръснати или комбинирани в големи плътни купове.
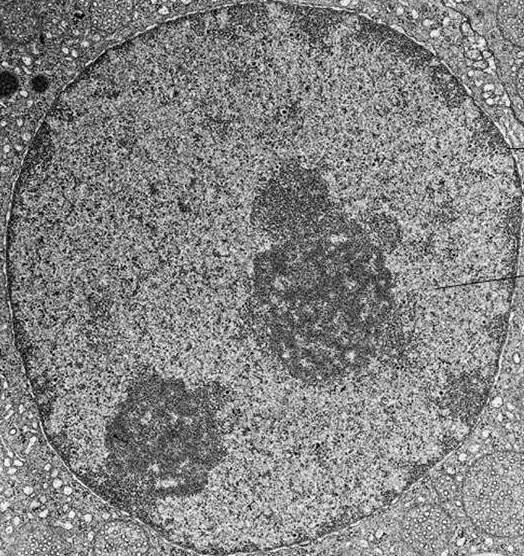
Хроматиновата кондензация по време на клетъчното делене води до образуването на хромозоми, които са видими дори в конвенционален светлинен микроскоп.
Структурни и функционални компоненти на хроматина
За да се определи какъв хроматин е на биохимично ниво, учените извличат това вещество от клетките, прехвърлят го в разтвор и изследват съставния състав и структурата на компонента в тази форма. Използвани са както химични, така и физични методи, включително технологии за електронна микроскопия. Оказа се, че химичният състав на хроматина с 40% е представен от дълги ДНК молекули и с почти 60% от различни протеини. Последните са разделени на две групи: хистони и нехистони.
Хистоните са голямо семейство ядрени белтъци, които се свързват силно с ДНК, образувайки структурен скелет на хроматина. Техният брой е приблизително равен на процента на генетичните молекули.
Останалите (до 20%) от протеиновата фракция попадат върху ДНК-свързващи и пространствено-модифициращи протеини, както и ензими, участващи в процеса на четене и копиране на генетична информация.
В допълнение към основните елементи, в състава на хроматиновите рибонуклеинови киселини (РНК), гликопротеините, въглехидратите и липидите се намират в малка част, но въпросът за тяхната връзка с комплекта ДНК опаковки е все още отворен.
Хистони и нуклеозоми
Молекулното тегло на хистоните варира от 11 до 21 kDa. Голям брой основни аминокиселинни лизинови и аргининови остатъци дават на тези протеини положителен заряд, допринасяйки за образуването на йонни връзки с противоположно заредени фосфатни групи на двойната двойна спирала на ДНК.
Има 5 вида хистони: H2A, H2B, H3, H4 и H1. Първите четири вида участват в образуването на основната структурна единица на хроматина - нуклеозома, който се състои от ядрото (протеиново ядро) и ДНК, обвита около нея.
Нуклеозомното ядро е представено от осем молекулно октамерен комплекс от хистони, който включва тетрамер НЗ-Н4 и димер Н2А-Н2В. Област от ДНК с дължина от около 146 нуклеотидни двойки се навива върху повърхността на протеиновата частица, образувайки 1.75 намотки, и преминава в линкерна последователност (приблизително 60 bp), свързваща нуклеозомите един с друг. H1 молекулата се свързва с линкерната ДНК, като я предпазва от действието на нуклеазите.
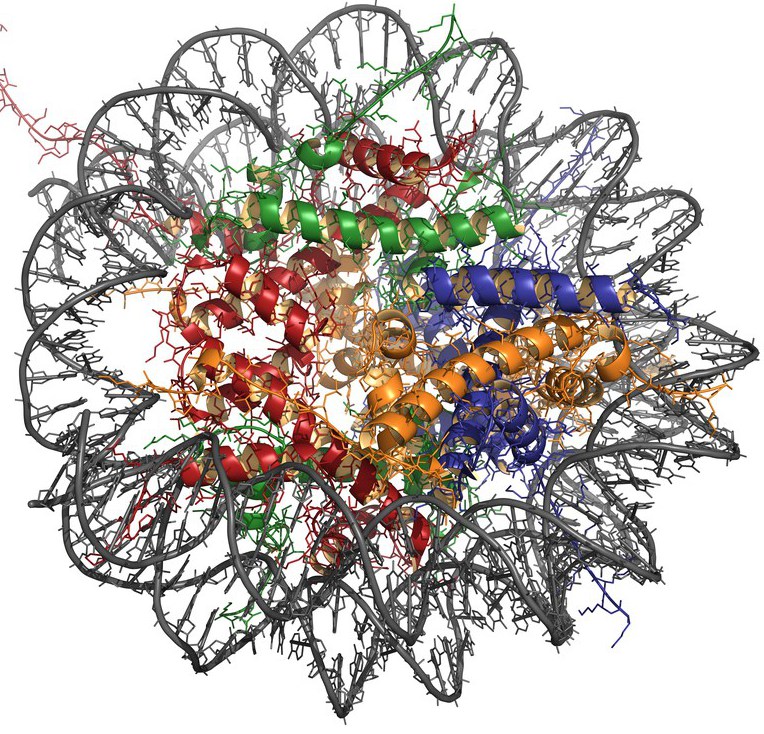
Хистоните могат да претърпят различни модификации, като ацетилиране, метилиране, фосфорилиране, ADP-рибозилиране и взаимодействие с убиквитинов протеин. Тези процеси влияят върху пространствената конфигурация и плътността на ДНК опаковките.
Нехистонови протеини
Има няколкостотин разновидности на нехистонови протеини с различни свойства и функции. Молекулното им тегло варира от 5 до 200 kDa. Специална група се състои от специфични за сайта протеини, всеки от които е допълващ към специфична област от ДНК. Тази група включва 2 семейства:
- "цинкови пръсти" - разпознават фрагменти от 5 нуклеотидни двойки;
- хомодимери се характеризират със структура на спирала-обратна спирала в фрагмент, свързан с ДНК.
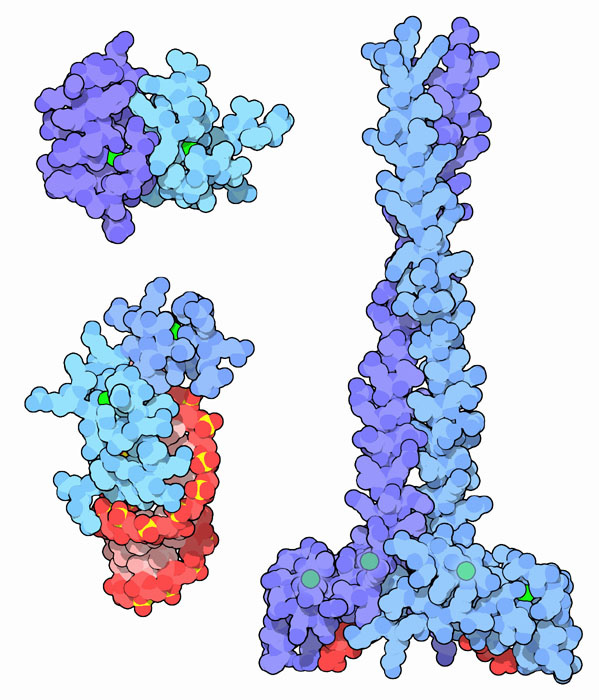
Най-добре проучени са така наречените протеини с висока мобилност (HGM протеини), които са постоянно свързани с хроматина. Това име на семейството получи поради високата скорост на движение на протеиновите молекули в електрофоретичния гел. Тази група заема по-голямата част от нехистонната фракция и включва четири основни вида протеини на HGM: HGM-1, HGM-14, HGM-17 и HMO-2. Те изпълняват структурни и регулаторни функции.
Нехистоновите протеини включват също ензими, които осигуряват транскрипция (синтез на информационна РНК), репликация (удвояване на ДНК) и възстановяване (възстановяване на увреждане в генетичната молекула).
Нива на уплътняване на ДНК
Особеността на структурата на хроматина е такава, че позволява ДНК нишките с обща дължина повече от един метър да се поберат в ядро с диаметър около 10 микрона. Това е възможно благодарение на многоетапна система за опаковане на генетични молекули. Общата схема на уплътняване включва пет нива:
- нуклеозомна нишка с диаметър 10–11 nm;
- фибрила 25–30 nm;
- домейни на линии (300 nm);
- Влакно с дебелина 700 nm;
- хромозоми (1200 nm).
Тази форма на организация осигурява намаляване на дължината на оригиналната ДНК молекула с 10 хиляди пъти.
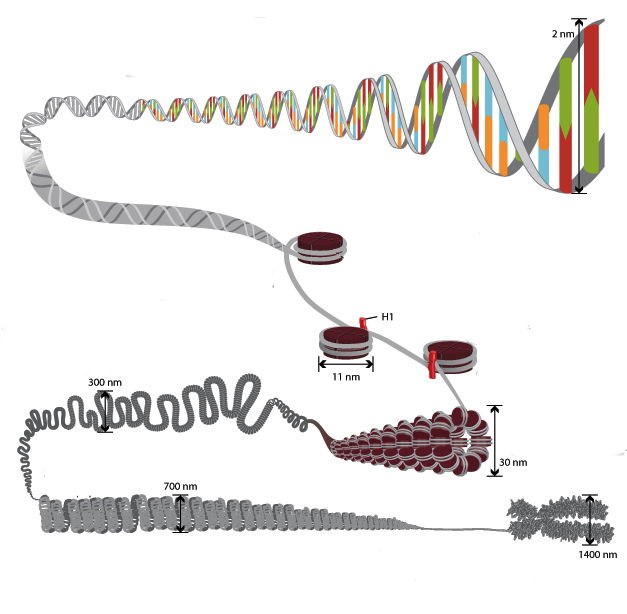
11 nm нишка се образува от серия от нуклеозоми, свързани чрез линкерни ДНК области. На електронни микрографии такава структура прилича на мъниста, нанизана на въдица. Нуклеозомната нишка се сгъва в бобина като соленоид, образувайки фибрила с дебелина от 30 nm. Хистон Н1 участва във формирането му.
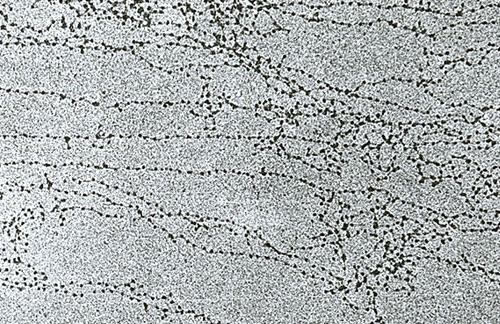
Соленоидната фибрила се сгъва в контури (известни още като области), които са фиксирани върху поддържащата интрануклеарна матрица. Всеки домейн съдържа от 30 до 100 хиляди двойки нуклеотиди. Това ниво на уплътняване е характерно за интерфазен хроматин.
Структурата с дебелина от 700 nm се образува чрез спиралообразуване на фибрилната област и се нарича хроматид. На свой ред, две хроматиди образуват петото ниво на ДНК организация - хромозома с диаметър 1400 nm, която става видима на етапа на митоза или мейоза.
Така хроматинът и хромозомата са форми на опаковане за генетичен материал, които зависят от жизнения цикъл на клетката.
хромозом
Хромозомата се състои от две сестрински хроматиди, идентични една на друга, всяка от които се формира от една суперспирали ДНК молекула. Половините са свързани чрез специално тяло на фибрила, наречено центромер. В същото време, тази структура е свиване, което разделя всяка хроматида върху раменете.
За разлика от хроматина, който е структурен материал, хромозомата е дискретна функционална единица, характеризираща се не само по структура и състав, но и с уникален генетичен набор, както и с определена роля в прилагането на механизмите на наследственост и вариабилност на клетъчното ниво.
Еухроматин и хетерохроматин
Хроматинът в ядрото съществува в две форми: по-малко спирализирано (еухроматин) и по-компактно (хетерохроматин). Първата форма съответства на транскрипционно активните секции на ДНК и следователно не е толкова строго структурирана. Хетерохроматинът се подразделя на незадължително (може да премине от активна към плътна неактивна форма в зависимост от етапа на жизнения цикъл на клетката и необходимостта от прилагане на определени гени) и от конституирането (постоянно кондензирано). По време на митотично или мейотично разделяне, целият хроматин е неактивен.
Конститутивният хетерохроматин е намерен близо до центромерите и в крайните области на хромозомата. Резултатите от електронната микроскопия показват, че такъв хроматин запазва висока степен на кондензация не само на етапа на клетъчното делене, но и по време на интерфазата.
Биологичната роля на хроматина
Основната функция на хроматина е гъсто опакована с голямо количество генетичен материал. Обаче, просто поставянето на ДНК в ядрото не е достатъчно, за да функционира клетката. Необходимо е тези молекули да "работят" правилно, т.е. те могат да прехвърлят информацията, съдържаща се в тях, чрез системата ДНК-РНК-протеин. В допълнение, клетката трябва да разпределя генетичен материал по време на разделянето.
Хроматиновото устройство напълно отговаря на тези цели. Протеиновата част съдържа всички необходими ензими и особеностите на структурата им позволяват да взаимодействат с определени участъци от ДНК. Следователно, втората важна функция на хроматина е да осигури всички процеси, свързани с прилагането на ядрения геном.