Комплекс Голджи: описание
Комплекс Голджи е открит през 1898 година. Тази мембранна структура е проектирана да отделя съединения, които се синтезират в ендоплазмения ретикулум. Освен това ще се запознаем с тази система. 
Комплекс Голджи: сграда
Устройството е купчина от мембранни дискове с формата на дискове. Тези торбички са донякъде удължени към краищата. Балонната система на Голджи е свързана с резервоарите. В животински клетки присъстват един голям или няколко пилоти, които са свързани чрез тръби, в растителни клетки Откриват се диктиозоми (няколко отделни купчини). Комплекс Голджи включва три отдела. Те са заобиколени от мембранни мехурчета:
- cis-най-близко до ядрото;
- медийната;
- транс-отдел (най-отдалечен от ядрото).
Тези системи се отличават с набор от ензими. В отдела за цис, първата торбичка се нарича "спасителен резервоар". С негова помощ, рецепторите, които идват от ендоплазмената междинна мрежа, се връщат назад. Ензимен цис отдел, наречен фосфогликозидаза. Добавя фосфат към маноза (въглехидрати). В медиалната част се намират два ензима. Това, по-специално, менадиаза и N-ацетилглюкозамин трансфераза. Последният добавя гликозамини. Ензими на транс-отдел: пептидаза (осъществява протеолиза) и трансфераза (с нейната помощ се осъществява трансферът на химични групи). 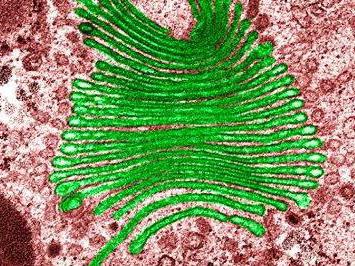
Комплекс Голджи: функции
Тази структура осигурява отделянето на протеини в следните три потока:
- Лизозомни. Според него, гликираните протеини проникват в цис-отдела на апарата на Голджи. Някои от тях са фосфорилирани. В резултат на това се образува маноза-6-фосфат - пазазозомални ензими. В бъдеще тези фосфатирани протеини ще влязат в лизозомите и няма да бъдат модифицирани.
- Конститутивна екзоцитоза (секреция). Този поток включва протеини и липиди, които са станали компоненти на апарата на клетъчната повърхност, включително гликокаликс. Също така, могат да присъстват съединения, които са част от извънклетъчния матрикс.
- Индуцирана секреция. Протеините, които функционират извън клетката, повърхностни апарати, проникват в този поток във вътрешната среда на тялото. Индуцираната секреция е характерна за секреторните клетки.
Комплекс Голджи участва в образуването на секреция на лигавици - мукополизахариди (гликозаминогликани). Устройството също така образува въглехидратни компоненти на гликокаликса. Те са предимно гликолипиди. Системата също така осигурява сулфатиране на протеинови и въглехидратни елементи. Комплексът Голджи участва в частичната протеолиза на протеините. В някои случаи, поради това, съединението от неактивното преминава в активна форма (например, проинсулинът се трансформира в инсулин). 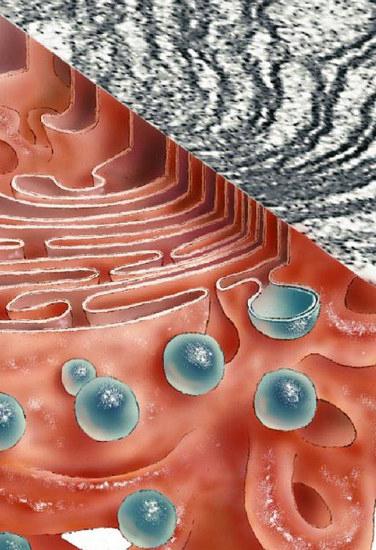
Преместване на съединения от ендоплазмения ретикулум (EPS)
Комплексът е асиметричен. По-близо до ядрото на клетката се включват най-незрелите протеини. Везикулите, мембранните везикули се прикрепят непрекъснато към тези торбички. Те се отделят от ендоплазмения гранулиран ретикулум. На неговите мембрани преминава процесът на протеинов синтез чрез рибозоми. Транспортирането на съединения от ендоплазмения ретикулум към комплекса Голджи е безразборно. В същото време неправилно или непълно сгънати протеини продължават да остават в EPS. Обратното движение на съединения в ендоплазмения ретикулум изисква специална сигнална последователност и е станало възможно благодарение на свързването на тези вещества към мембранните рецептори в цис-отделението.
Протеинова модификация
В резервоарите на комплексното узряване настъпват съединения, които са предназначени за секреция, трансмембрани, лизозоми и други вещества. Тези протеини последователно се движат по танкове към органелите. Техните модификации започват в тях - фосфолиране и гликозилиране. По време на първия процес, остатъкът се прикрепя към протеините. фосфорна киселина. В О-гликозилирането, сложните захари се кукират през кислородния атом. Различните резервоари съдържат различни каталитични ензими. Следователно, последователно протичат различни процеси със зреещи в тях протеини. Несъмнено подобно стъпало трябва да се контролира. Като вид "марка за качество" се използват полизахаридни остатъци (главно маноза). Те обозначават зреещи протеини. По-нататъшното движение през резервоарите на съединенията не е напълно разбрано от науката, въпреки факта, че устойчивите вещества остават по-малко или повече свързани с една торба. 
Транспортиране на протеини от апарата
От междинните мехурчета. Те съдържат напълно зрели протеинови съединения. Основната функция на комплекса е сортирането на протеини, преминаващи през него. В апарата се образува "трипосочен протеинов поток" - съзряване и транспорт:
- съединения плазмена мембрана.
- Как да стигнем.
- Лизозомни ензими.
Чрез везикуларен транспорт, протеини, преминали през комплекса Голджи, се доставят на определени места в съответствие с "етикетите". Този процес също не е напълно разбран от науката. Установено е, че транспортирането на протеини от комплекса изисква участието на специфични мембранни рецептори. Те разпознават съединението и осигуряват селективно скачване на везикула и една или друга органела. 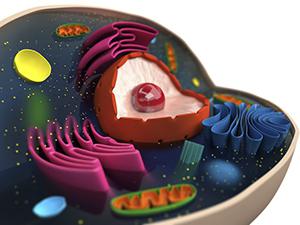
Образуване на лизозоми
Чрез апарата преминава много хидролитични ензими. Добавянето на посочения по-горе етикет се извършва с участието на два ензима. Специфичното разпознаване на лизозомните хидролази от елементите на тяхната третична структура и добавянето на N-ацетилглюкозамин фосфат се извършва от N-ацетилглюкозамин фосфотрансферазата. Фосфогликозидът, вторият ензим, отстранява N-ацетилглюкозамина, което води до образуването на M6F етикет. Той, от своя страна, се разпознава от рецепторния протеин. С негова помощ, хидролазите се доставят до везикулите и техните мандати в лизозомите. В кисели условия, фосфатът се разцепва от зрялата хидролаза в тях. Ако има нарушения в активността на N-ацетилглюкозамин фосфотрансфераза поради мутации или поради генетични дефекти в M6F рецептора, всички лизозомни ензими се доставят по подразбиране към външната мембрана. След това те се секретират в екстрацелуларни условия. Установено е също, че някои от M6P рецепторите също са транспортирани до външната мембрана. Те извършват връщане на случайно уловени лизозомни ензими от външната среда вътре в клетката по време на ендоцитоза. 
Транспортиране на вещества към външната мембрана
Обикновено, дори на етапа на синтеза, протеиновите съединения на външната мембрана с техните хидрофобни области са вградени в стената на ендоплазмения ретикулум. След това се доставят в комплекса Голджи. Оттам те се транспортират до клетъчната повърхност. В процеса на сливане на плазмалемата и везикулите, такива съединения не се освобождават в околната среда.
секреция
Почти всички съединения, произведени в клетката (както от протеинова, така и от не-протеинова природа) преминават през комплекса Голджи. Там те се развиват в секреторни везикули. Така при растенията с диктофони материалът се произвежда. клетъчна стена.